GYMNOSPERMAE
A. PENGERTIAN
Tumbuhan dibagi menjadi dua, yaitu tumbuhan tak berpembuluh (non vaskuler) dan tumbuhan berpembuluh (vaskuler). Tumbuhan tak berpembuluh yaitu lumut, sedangkan tumbuhan berpembuluh terdiri atas tumbuhan tak berbiji, yaitu paku dan tumbuhan berbiji. Sedangkan tumbuhan berbiji sendiri dibagi dalam tumbuhan berbiji terbuka (gymnospermae) dan tumbuhan berbiji tertutup (angiospermae).
Tumbuhan berbiji ( Spermatophyta ) adalah tumbuhan yang mempunyai bagian yang di sebut biji. Pada dasarnya tumbuhan biji itu dicirikan dengan adanya bunga sehingga sering disebut dengan tumbuhan berbunga (Anthopyta). Biji dihasilkan oleh bunga setelah terjadi peristiwa penyerbukan dan pembuahan. Dengan kata lain, biji dapat dihasilkan merupakan alat pembiakan secara seksual (generatif). Selain itu, ada juga pembiakan secara aseksual (vegetatif).
Tumbuhan berbiji di kelompokkan menjadi dua anak divisi, yaitu tumbuhan berbiji terbuka (Gymnospermae) dan tumbuhan biji tertutup (Angiospermae). Pada tumbuhan biji terbuka, biji tertutup dengan daging buah atau daun buah (karpelum). Misalnya, pada cemara, pinus, dan damar. Sementara itu, pada tumbuhan berbiji tertutup, biji di tutupi oleh daging buah atau daun buah. Misalnya, pada mangga, durian, dan jeruk. Dalam tumbuhan berbiji banyak sekali ordo ataupuun famili dari tiap divisi. Hal ini membuktikan bahwa tumbuhan berbiji merupakan tumbuhan yang dapat dikatakan tumbuhan yang memiliki bagian yang sangatlah banyak.
Gymnospermae adalah tumbuhan yang memiliki biji terbuka. Gymnospermae berasal dari bahasa Yunani, yaitu gymnos yang berarti telanjang dan sperma yang berarti biji, sehingga gymnospermae dapat diartikan sebagai tumbuhan berbiji terbuka.tumbuhan berbiji terbuka merupakan kelompok tumbuhan berbiji yang bijinya tidak terlindung dalam bakal buah (ovarium). Secara harfiah Gymnospermae berarti gym = telanjang dan spermae = tumbuhan yang menghasilkan biji. Pada tumbuhan berbunga (Angiospermae atau Magnoliphyta), biji atau bakal biji selalu terlindungi penuh oleh bakal buah sehingga tidak terlihat dari luar. Pada Gymnospermae, biji nampak (terekspos) langsung atau terletak di antara daun-daun penyusun strobilus atau runjung.
Gymnospermae telah hidup di bumi sejak periode Devon (410-360 juta tahun yang lalu), sebelum era dinosaurus. Pada saat itu, Gymnospermae banyak diwakili oleh kelompok yang sekarang sudah punah dan kini menjadi batu bara : Pteridospermophyta (paku biji), Bennettophyta dan Cordaitophyta. Anggota-anggotanya yang lain dapat melanjutkan keturunannya hingga sekarang. Angiospermae yang ditemui sekarang dianggap sebagai penerus dari salah satu kelompok Gymnospermae purba yang telah punah (paku biji).
Gymnospermae berasal dari Progymnospermae melalui proses evolusi biji. Hal tersebut dapat dilihat dari bukti-bukti morfologi yang ada. Selanjutnya Progymnospermae dianggap sebagai nenek moyang dari tumbuhan biji. Progymnospermae mempunyai karakteristik yang merupakan bentuk antara Trimerophyta dan tumbuhan berbiji. Meskipun kelompok ini menghasilkan spora, tetapi juga menghasilkan pertumbuhan xylem dan floem sekunder seperti pada Gymnospermae. Progymnospermae juga sudah mempunyai kambium berpembuluh yang bifasial yang mampu menghasilkan xilem dan floem sekunder. Kambium berpembuluh merupakan ciri khas dari tumbuhan berbiji. Salah satu contoh Progymnospermae adalah tipe Aneurophyton yang hidup pada jaman Devon, sudah menunjukkan system percabangan tiga dimensi dengan stelenya yang bertipe protostele. Contoh lainnya adalah tipe Archaeopteris yang juga hidup di jaman Devon. Kelompok ini dianggap lebih maju karena sudah menunjukkan adanya system percabangan lateral yang memipih pada satu bidang dan sudah mempunyai struktur yang dianggap sebagai daun. Batangnya mempunyai stele yang bertipe eustele yang menunjukkan adanya kekerabatan dengan tumbuhan berbiji yang sekarang.
B. KLASIFIKASI
Gymnospermae terdiri dari beberapa divisi baik yang sudah punah maupun yang masih ada sampai sekarang, yaitu mencakup 3 divisi yang telah punah dan 4 divisi yang masih bertahan.
Tumbuhan Gymnospermae yang sudah punah
Tiga divisi yang sudah punah adalah:
Bennetophyta
Cordaitophyta
Pteridospermophyta, sudah punah namun dianggap sebagai moyang Angiospermae.
Divisi Cycadophyta, yang mempunyai daun menyerupai palem, agak menyerupai tumbuhan Cycas yang sekarang. Kelompok ini (Bennetitales) juga mengikuti garis evolusi yang sama seperti tumbuhan berbiji yang ada sampai sekarang. Namun terdapat perbedaan, yaitu sifat biseksualisme pada strobilusnya dan aspek lainnya. Kelompok yang menyerupai Cycas ini hidup pada jaman Jura dan Creta.
Tumbuhan Gymnospermae yang masih ada sampai sekarang
Empat divisi Gymnospermae yang masih bertahan adalah:
1) Ginkgophyta
Hanya satu jenis yang masih bertahan: Ginkgo biloba
2) Cycadophyta
Cycadophyta di bagi menjadi dua famili, yaitu Cycadaceae dan Zamiaceae.
3) Coniferophyta atau dapat disubut Pinophyta
Merupakan tumbuhan runjung.
4) Gnetophyta
Dengan anggota hanya 3 genus: Gnetum(melinjo dan kerabatnya), Welwitschia, dan Ephendra.
C. HABITAT
Gymnospermae hidup di mana-mana, hampir di seluruh permukaan bumi ini. Mulai dari daerah tropis hingga daerah kutub dan dari daerah yang cukup air hingga daerah kering.
Ginkgophyta
Banyak ditemukan di negara Cina, khususnya di daerah kecil di Zhejiang Cina dan di Mu Tian Shan. provinsi di Timur
Cycadophyta
Cycadophyta hidup di daerah tropis dan subtropis.
Coniferophyta atau dapat disubut Pinophyta
Tumbuhan yang termasuk Coniferophyta hidup tersebar di berbagai daerah, bahkan hampir di seluruh daerah di dunia. Pohon pinus dan cemara banyak tumbuh di Eropa bagian pegunungan.
Gnetophyta
Banyak tumbuh di daerah tropis dan subtropis.
D. CIRI-CIRI
Gymnospermae memiliki ciri-ciri sebagai berikut:
1. Bakal biji tidak terlindungi oleh daun buah.
2. Pada umumnya perdu atau pohon, tidak ada yang berupa herba. Batang dan akar berkambium sehingga dapat tumbuh membesar. Akar dan batang tersebut selalu mengadakan pertumbuhan menebal sekunder. Berkas pembuluh pengangkutan kolateral terbuka. Xilem pada gymnospermae hanya terdiri atas trakeid saja sedangkan floemnya tanpa sel-sel pengiring.
3. Mempunyai akar, batang, dan daun sejati.
4. Bentuk perakaran tunggang.
5. Daun sempit, tebal dan kaku.
6. Tulang daun tidak beraneka ragam.
7. Tidak memiliki bunga sejati.
8. Alat kelamin terpisah, serbuk sari terdapat dalam strobilus jantan dan sel telur terdapat dalam strobilus betina.
9. Struktur perkembangbiakan yang khas adalah biji yang dihasilkan bunga ataupun runjung. Setiap biji mengandung bakal tumbuhan , yaitu embrio yang terbentuk oleh suatu proses reproduksi seksual. Sesudah bertunas embrio ini tumbuh menjadi tumbuhan dewasa.
10. Sperma atau sel kelamin jantan menuju kesel telur atau sel kelamin betina melalui tabung serbuk sari hanya terdapat pada tumbuhan berbiji.
11. Tumbuhan biji mempunyai jaringan pembuluh yang rumit. Jaringan ini merupakan saluran menghantar untuk mengangkut air, mineral, makanan dan bahan – bahan lain.
12. Tumbuhan berbiji terbuka memiliki pigmen hijau (klorofil) yang penting untuk fotosintesis yaitu suatu proses dasar pembuatan makanan pada tumbuhan.
13. Gymnospermae memiliki batang yang tegak lurus dan bercabang-cabang. Daunnya jarang yang berdaun lebar, jarang yang bersifat majemuk, dan system pertulangan daunnya tidak banyak ragamnya. Hal ini sangat berbeda dengan karakteristik daun yang terdapat pada angiospermae yang sistem pertulangannya beraneka ragam.
E. REPRODUKSI
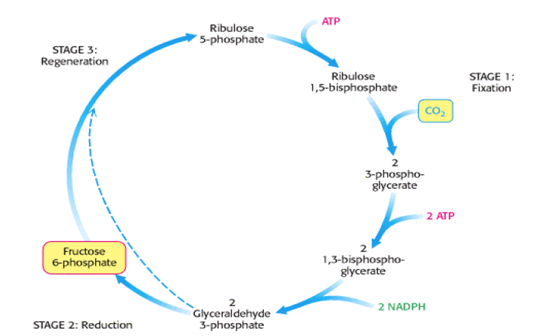
Organ reproduksi pada gymnospermae disebut konus atau strobilus. Tumbuhan berbiji terbuka tidak memiliki bunga, sporofil terpisah-pisah atau membentuk srobilus jantan dan betina. Makrosporofil dan makrosporangium yang tampak menempel pada strobilus betina. Letak makrosporofil dan mikrosporofil terpisah. Sel kelamin jantan berupa spermatozoid yang masih bergerak aktif. Di dalam strobilus jantan terdapat banyak anteridium yang mengandung sel-sel induk butir serbuk. Sel-sel tersebut bermeiosis dari setiap sel induk terbentuk 4 butir serbuk yang bersayap. Pada strobilus betina terdapat banyak arkegonium. Pada tiap-tiap arkegonium terdapat satu sel induk lembaga yang bermeiosis sehingga terbentuk 4 sel yang haploid. Tiga mati, dan satu sel hidup sebagai sel telur. Arkegonium ini bermuara pada satu ruang arkegonium.
Pada Gymnospermae sering terjadi poliembrioni, walaupun hanya ada satu embrio yang terus berkembang karena adanya pembelahan beberapa arkegonia. Air sudah tidak digunakan sebagai media fertilisasi karena adanya pembentukan buluh serbuk pada serbuk sari yang berkecambah.
Pada Coniferophyta dan Gnetophyta spermanya tidak mempunyai flagel, sehingga buluh serbuk menghantarkannya langsung ke mulut arkegonia. Serta pada Cycas dan Gingko fertilisasinya merupakan bentuk antara kondisi pada paku-pakuan dan tumbuhan tanpa biji lainnya, yaitu spermanya mampu berenang bebas dan bentuk pada tumbuhan berbiji yaitu spermanya tidak mampu bergerak bebas.
Gametofi jantan umumnya bersifat haustorial, yaitu menyerap makanan dari ovulum ketika tumbuh, walaupun dibutuhkan buluh serbuk tetapi tidak langsung masuk ke arkegonium. Buluh serbuk tersebut tumbuh dan menetap di dalam nuselus selama berbulan-bulan sebelum menuju gametofit betina. Setelah sampai di mulut gametofit betina, buluh serbuk robek dan melepaskan sel sperma yang berflagel banyak. Sperma tersebut kemudian menuju ke arkegonium dan membuahi telur. Dengan adanya buluh sperma tersebut maka tumbuhan berbiji tidak ada lagi yang bergantung pada ketersediaan air pada fertilisasinya.
Proses Penyerbukan dan Pembuahan
Penyerbukan yang terjadi pada tumbuhan berbiji terbuka selalu dengan cara anemogami (penyerbukan dengan bantuan angin). Serbuk sari jatuh langsung pada bakal biji. Selang waktu antara penyerbukan sampai pembuahan relatif panjang. Pembuahan yang terjadi pada gymnospermae disebut pembuahan tunggal (setiap inti generatif melebur dengan inti sel telur). Mikropil terdedah ke udara bebas. Pembuahan pada gymnospermae disebut pembuahan tunggal, karena tiap-tiap inti sperma membuahi satu sel telur.